Nachts sind alle Nebel grau
Eine astronomische Sicht auf die Physiologie des Sehens

| Grundlegendes | Dunkeladaption | Stäbchen vs. Zapfen |
|
Nachts sind alle Nebel grau Eine astronomische Sicht auf die Physiologie des Sehens |
|
Teil 2
Dunkeladaption: Was passiert in unserem Auge, wenn es dunkel wird?
| Eine der beeindruckendsten Eigenschaften unseres Auges besteht in seiner Fähigkeit sich an die unterschiedlichsten Lichtintensitäten in unserer Umgebung anpassen zu können. In diesem zweiten Teil möchte ich ein wenig auf die dahinter liegenden Mechanismen eingehen. |  |
 |
Lichtadaption Lichtadaption beschreibt die Mechanismen, mit Hilfe derer das Auge seine Empfindlichkeit herunter reguliert, um sich an helles Umgebungslicht anzupassen. |
Bei der Lichtadaption
unterscheidet man zwei Mechanismen, die sich grundlegend unterscheiden:
Durch diese Anpassung wird unser Auge in die Lage versetzt, Lichtstimuli über einen Intensitätsbereich von 1 000 000 000 unterscheiden zu können. |
|
| Dunkeladaption Bei der Dunkeladaption laufen die oben genannten Schritte in umgekehrter Richtung ab:
Warum ist dieser Schritt bei der Dunkeladaption so viel langsamer?
|
 |
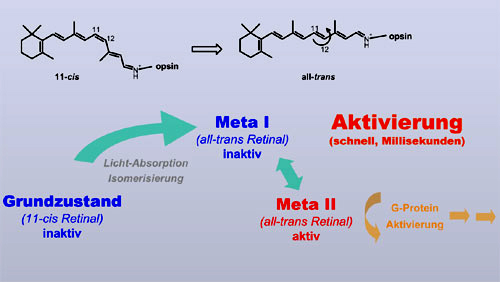 |
Aktivierung und Deaktivierung
von Rhodopsin Um dies verstehen zu können, müssen wir uns die Aktivierung und nachfolgende Deaktivierung von Rhodopsin genauer anschauen. Die Aktivierung von Rhodopsin wird, wie im ersten Teil erläutert, durch die Photoisomerisierung des Retinals von 11-cis zu all-trans getriggert. Der Rezeptor reagiert darauf und passt sich an den isomerisierten Liganden in einer Folge von inaktiven Intermediaten an. Erst im Übergang vom noch inaktiven Meta-I-Zustand zum aktiven Meta-II-Zustand nimmt er dann die für die G Protein-Aktivierung notwendige Konformation ein. Diese Aktivierungsschritte erfolgen sehr schnell, auf der Zeitskala von Millisekunden. |
 |
Das Rezeptorsignal muss auch sehr
schnell wieder abgeschaltet werden, um einen Sehprozess mit ausreichender
Zeitauflösung zu gewährleisten. Dies erfolgt durch rasche Bindung von
Kinasen und Arrestin an den aktivierten Rezeptor, was die Aktivierung von G
Protein unterbindet. Die aktive Rezeptorkonformation von Meta II bleibt
dabei jedoch erhalten.
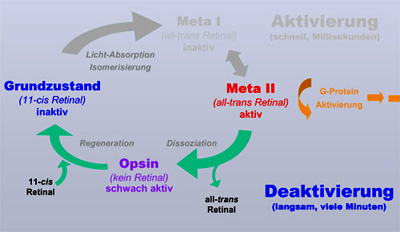
Sie zerfällt erst sehr viel langsamer, auf der Zeitskala von Minuten. In diesem Prozess wird die Retinal-Schiff-Base hydrolysiert und das all-trans Retinal verlässt die Bindungstasche. Übrig bleibt das Apoprotein Opsin, also das Rezeptorprotein ohne Retinal. Dieses kann dann, wieder auf der Zeitskala von Minuten, neu gebildetes 11-cis-Retinal binden und in den absolut inaktiven Grundzustand zurück kehren (Regeneration des Dunkelzustands). Der Zyklus ist somit geschlossen. |
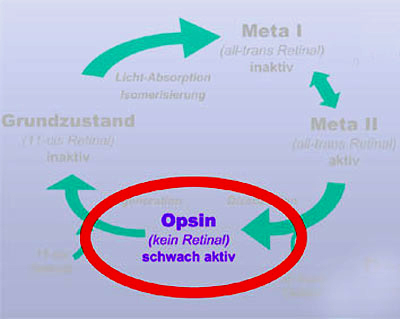
| Opsin ist schwach aktiv und
verhindert die Dunkeladaption So weit, so gut. 2001 konnte ich mit Hilfe von FTIR-Differenzenspektroskopie zeigen, dass Opsin ein Konformationsgleichgewicht einnimmt zwischen einer inaktiven und einer aktiven Rezeptor-Konformation (Vogel et al. (2001) JBC 276:38487). Dieses Konformations-Gleichgewicht liegt fast vollständig auf der Seite der inaktiven Konformation, aber eben nur fast. Die resultierende schwache Aktivität des Opsins ist ausreichend, um der Photorezeptorzelle "Licht" vor zu gaukeln, auch bei vollständiger Dunkelheit. Erst bei letztendlicher Bindung von 11-cis-Retinal und dem Übergang in den Dunkelzustand wird diese Aktivität nach Minuten dann vollständig abgeschaltet. Solange also noch Opsin aus vorhergehenden Aktivierungen vorhanden ist, "sieht" die Zelle virtuelles Licht und macht keine vollständige Dunkeladaption. Ein häufig auftretendes Missverständnis ist, dass die Empfindlichkeits-Steigerung bei der Dunkeladaption durch die Zunahme der Menge an Rhodopsin (durch Regeneration des Dunkelzustands) verursacht wird. Diese bewegt sich jedoch bei den für uns relevanten Beleuchtungsstärken im Bereich <<1% und ist somit nicht relevant (die Empfindlichkeitssteigerung kann mehrere Größenordnungen umfassen!) |
 Vogel et al. (2001) JBC 276:38487
|
|
Dunkeladaption Zusammenfassen kann man also sagen:
|
| Warum Rotlicht beim Beobachten? Beim Beobachten verwenden wir Rotlicht (sollten wir jedenfalls!), um unsere Dunkeladaption nicht zu beinträchtigen. Dies bezieht sich jedoch nur auf die Stäbchen, da das Rotlicht die rotempfindlichen Zapfen durchaus aktiviert (und dies ja auch tun soll, wir wollen mit diesen Zapfen ja schließlich Karten lesen). Damit die Stäbchen durch das Rotlicht möglichst wenig beeinträchtigt werden, muss die Wellenlänge des Lichts so gewählt werden, dass das Stäbchen-Rhodopsin nicht oder zumindest so wenig wie möglich aktiviert wird. Dies gelingt am besten im tiefroten Bereich über 630 nm. |
 |
| Durch das Rotlicht erreichen wir,
dass die Stäbchenzellen weitestgehend dunkeladaptiert bleiben. Dafür nehmen
wir in Kauf, dass die roten L-Zapfen durch das Rotlicht immer wieder
lichtadaptieren und nicht mehr dunkeladaptieren können. Dies stört uns
jedoch weniger, weil nachts
sowieso nur die Stäbchen für die Beobachtung von Deep Sky Objekten
empfindlich genug sind.
Aber warum ist das so? Darum geht es im nächsten Teil. |
weiter zu Stäbchen vs. Zapfen: Warum sehen wir nachts nur mit den Stäbchen gut?
![]()
| Grundlegendes | Dunkeladaption | Stäbchen vs. Zapfen |