All cats are gray in the dark
An astronomical view on the physiology of vision

| rhodopsin activation | dark adaptation | rods vs. cones |
|
All cats are gray in the dark An astronomical view on the physiology of vision |
|
part 2
Dark adaptation: What happens in our eye when it's getting dark?
| One of the most
impressive achievements of our eye is its ability to adapt to the most
different light intensities of our environment. In this second part, I will
do into the details involved in these adaptation processes.
|
 |
 |
Light adaptation Light adaptation describes the mechanism by which our eye reduces its sensitivity to light to adapt to a brightly illuminated environment. |
Light adaptation
involves (at least) two entirely different mechanisms:
This adaptation process enables our eye to differentiate between light stimuli over the vast intensity range of 1 000 000 000. |
|
| Dark adaptation During dark adaptation, the steps mentioned above are reversed:
Why is this last step so much slower during dark adaptation as compared with light adaptation?
|
 |
 |
Activation and
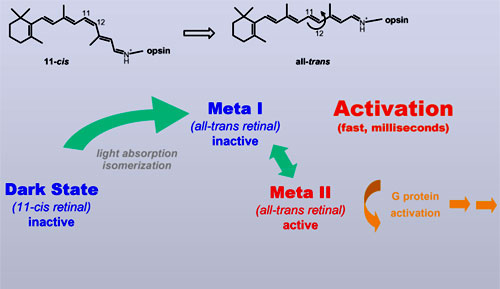
deactivation of rhodopsin In order to answer this question, we need to go deeper into the activation and the subsequent deactivation process of rhodopsin. Activation of rhodopsin is, as detailed in part one, triggered by the light-induced isomerization of retinal from 11-cis to all-trans. The receptor protein responds to this altered shape of the isomerized ligand via a series of (still inactive) intermediates. Only in the final transition form the inactive Meta I state to the active Meta II state the receptor adopts the conformation that finally activates the G protein. These activating steps occur very fast, on the time scale of several milliseconds.
|
 |
Once activated, the
receptor signal has to be turned off sufficiently fast to warrant a
reasonable time resolution (imagine watching an action movie with a time
resolution of our visual perception of, say, 10 seconds. Boring!). This is
achieved by rapid binding or rhodopsin kinase and arrestin to the activated
receptor, which interrupts binding and hence activation of G protein. The
active Meta II receptor conformation, however, is not altered by these
steps.
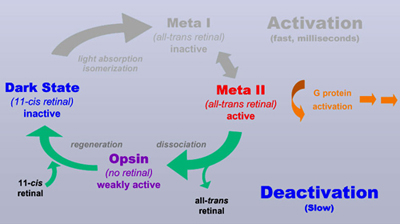
The Meta II conformation decays on a much slower time scale, involving minutes. During this process, the retinal Schiff base is hydrolyzed and all-trans retinal is released from its binding pocket. What's left is the receptor protein without retinal, which is termed opsin. Opsin will eventually bind newly synthesized 11-cis retinal and return by this process to the dark state, closing the visual cycle. This last step, however, takes again several minutes.
|
| Opsin is weakly
active and prevents complete dark adaptation So what's the point of this all? In 2001 I could show, using FTIR difference spectroscopy, that opsin may adopt two different conformations, an active one and an inactive one, both forming a conformational equilibrium (Vogel et al. (2001) JBC 276:38487). This conformational equilibrium is under physiological conditions almost fully on the side of the inactive conformation. Almost fully, but not completely. A tiny fraction of opsin adopts hence its active conformation. This tiny fraction is sufficient to make the photoreceptor cell "see" light, even in complete darkness. Only upon subsequent binding of 11-cis retinal and regeneration of the dark state (taking many minutes), this activity is eventually turned off. As long as there is any opsin left from previous bright light situations, the cell persists to "see" virtual light and will not undergo dark adaptation. A quite common misconception attributes the increase of sensitivity during dark adaptation to the increase of rhodopsin (by regeneration of the dark state after bleaching). However, this increase is in the range <<1% for the illumination intensities that we encounter during observing and hence not really relevant for the increase in sensitivity (which during dark adaptation may involve several orders of magnitude!).
|
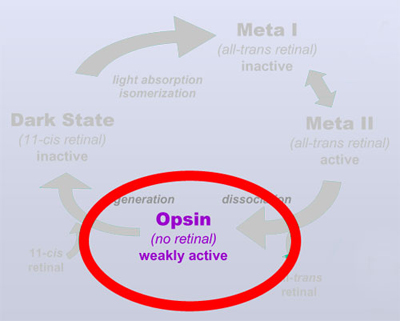 Vogel et al. (2001) JBC 276:38487
|
|
Dark adaptation Let's summarize this:
|
| What's the
point about red
light at the telescope? At the telescope, we use red light for reading charts (at least we should!), to keep our photoreceptor cells dark adapted. This is true only for our rod cells, as the red-sensitive L cones will be activated by red light (and should be, as we want to read our charts). In order to impair rod function as little as possible, we should choose a red light that activates rod rhodopsin not at all or at least as little as possible. This is best achieved by deep red light in the range above 630 nm.
|
 |
| By using red light,
we keep our rod cells mostly dark adapted. We put up with our red L cones
being activated by the red light and being light adapted again and again,
preventing any persistent dark adaptation. This is OK for us, as at night,
we use our rod cells only for observing deep sky objects. The cones are
simply not sensitive enough. But why aren't they? More about this in the following part.
|
read on Rods vs. cones: Why do we need our rod cells at night?
![]()
| rhodopsin activation | dark adaptation | rods vs. cones |